

Сотрудники лаборатории филогенетики и биохронологии Института экологии растений и животных (ИЭРиЖ УрО РАН) (г. Екатеринбург) ведут работы в области реконструкции истории современного биоразнообразия фауны Северной Евразии на генетическом, видовом и экосистемном уровнях и исследуют экологические основы эволюционных процессов, используя комплексный подход к изучению современных и вымерших организмов. Наблюдение за трансформацией экосистем и их отдельных компонентов в хронологическом срезе поможет выработать эффективные методы мониторинга и стратегии охраны видов континентальной фауны.
Современное биологическое разнообразие – это результат исторического развития органического мира, который может быть рассмотрен в различных масштабах времени. В актуальном времени разнообразие можно описать с необходимой степенью подробности, в историческом – восстановить его изменения по данным исторических документов, но лишь геологический масштаб времени позволяет установить, когда и при каких условиях появились современные виды и как выглядели экосистемы до того, как человек начал их активно осваивать. Реконструировать историю формирования существующего биологического разнообразия в масштабе геологического времени можно по данным палеонтологической летописи – остаткам животных и растений и следам их жизнедеятельности, которые сохранились в геологических отложениях. Но и ныне живущие организмы «помнят» тот путь, который прошли их предки в ходе своего эволюционного развития. Эта память сохраняется в таких изменчивых структурах организма, как генотип и фенотип.
Уральские ученые восстанавливают хронологию этапов формирования биологического разнообразия фауны центральной части Северной Евразии на генетическом, видовом и экосистемном уровнях, используя комплексный подход, сочетающий возможности изучения и сопоставления палеонтологических летописей и информации, которая содержится в генетических и фенотипических признаках современных видов, в особенностях их экологии.
Заведующий лабораторией филогенетики и биохронологии Института экологии растений и животных (ИЭРиЖ УрО РАН) (г. Екатеринбург), доктор биологических наук Александр Васильевич Бородин и сотрудники лаборатории – старший научный сотрудник, кандидат биологических наук Евгения Анатольевна Маркова и старший научный сотрудник, кандидат биологических наук Лидия Эдуардовна Ялковская – рассказали о деятельности лаборатории, фундаментальных и прикладных исследованиях, проходящих в рамках проектов РФФИ, в частности: что составляет основу подхода к реконструкции истории современного биоразнообразия и как можно определить возраст фауны, а также, как учет генетических данных раскрывает особенности внутривидового разнообразия и историю видов; как влияет на генетическое и фенотипическое разнообразие прохождение популяций через «бутылочное горлышко» и как об этом предупреждают признаки зубной системы.
«Появление лаборатории является, на мой взгляд, логическим продолжением развития экологической уральской школы, основу которой заложил Станислав Семёнович Шварц, и «отцы-основатели» Института экологии растений и животных, среди которых, в данном случае, нужно отметить и Н.В. Тимофеева-Ресовского. Тестирование эволюционных концепций (в том числе, и эволюционно-экологических) на макроэволюционном уровне возможно только с привлечением ископаемого материала. Как это ни парадоксально, но раньше в Институте экологии растений и животных такие исследования начали ботаники. Но появление палеозоологов в стенах ИЭРиЖ было неизбежно. Первым был Николай Георгиевич Смирнов, который привел меня (ещё студента) в лабораторию экологических основ изменчивости организмов Владимира Николаевича Большакова, где в это время уже работала ученица Тимофеева-Ресовского Эмилия Абрамовна Гилева. В дальнейшем Н.Г. Смирнов возглавил лабораторию исторической экологии (в которой работал и я), аккумулировавшей лучшие традиции и подходы четвертичной зоологии питерской школы Игоря Михайловича Громова и комплексных исследований в области исторической экологии московской школы Льва Георгиевича Динесмана. Эмилия Абрамовна развивала эколого-генетические методы изучения популяций млекопитающих, всегда находясь на передовых позициях и постановки задач исследований, и используемых методов, но она никогда не теряла интерес и к вопросам закономерностей морфологической эволюции. Когда в ее группе популяционной цитогенетики в конце 80-х годов начали использовать молекулярно-генетические методы, у нас с ней возникла идея попытаться сопоставить на некоторых модельных видах мелких млекопитающих степень морфологической и генетической дифференциации. Этот момент можно считать отправным в формировании наших совместных научных изысканий.
За годы сотрудничества с Эмилией Абрамовной и группой популяционной цитогенетики помимо ряда статей нами было реализовано несколько проектов РФФИ, в ходе выполнения которых оттачивалось взаимодействие и совершенствовались методики, используемые в эволюционно-палеоэкологических и эколого-генетических исследованиях. Итогом этого сотрудничества, собственно говоря, и является создание 1 апреля 2012 года лаборатории филогенетики и биохронологии. В состав новой лаборатории вошли сотрудники, ранее работавшие в лаборатории исторической экологии в группе под моим руководством, и сотрудники группы популяционной цитогенетики, которой на тот момент руководила Лидия Эдуардовна Ялковская. К сожалению, Эмилия Абрамовна в 2008 году ушла из жизни.
Сейчас в состав лаборатории входят как уже состоявшиеся ученые, ведущие исследования на мировом уровне, так и молодые сотрудники, только начинающие свой научный путь. Генетические исследования и работы в области экспериментальной экологии, научные идеи которых зародились еще во времена нашего сотрудничества с Э.А. Гилевой, успешно продолжают Л.Э. Ялковская, М.И. Чепраков, С.Б. Ракитин. Исследования в области палеозоологии, палео- и эволюционной экологии – Е.А. Маркова, Е.В. Зиновьев, А.Е. Некрасов, С.В. Зыков, М.А. Фоминых. Появление в составе лаборатории молодых специалистов – П.А. Сибирякова, Л.И. Сибиряковой, М.А. Крохалевой, а также недавних студентов и выпускников - А.Т. Галимова, А.О. Усольцевой и Д.С. Пилевич позволяет надеяться, что сформировавшиеся традиции не только будут поддержаны, но и будут развиваться в дальнейшем, – Александр Бородин сообщил об истоках зарождения лаборатории, её кадровом научном потенциале и о разработке новой методологической основы. –
Любое научное знание предполагает его использование для получения некой прогностической модели. Методологическая основа наших исследований отражается в сентенции «the present is the key to the past is the key to the future» (Mathieson, Elizabeth Lincoln (13 May 2002). Как я уже говорил, единственным прямым свидетельством реальности макроэволюционного процесса является палеонтологическая летопись. Но в свое время, академик Леонид Петрович Татаринов отмечал, что, если генетики, изучающие современные организмы, с уверенностью могут сказать, что у объектов их исследований были родители, то никто из палеонтологов не может утверждать, что организм, ископаемые остатки которого были обнаружены, кого-нибудь породил на свет. Палеонтолог может лишь утверждать, что в определённые моменты геологической истории на данных территориях жили такие-то существа с такими-то морфологическими характеристиками. Выстроить из этой последовательности палеонтологической летописи, зафиксированной в дискретных геологических слоях, эволюционную последовательность, связать ее с современным генетическим разнообразием и морфо-экологическими характеристиками современных животных – собственно говоря, на это направлена работа лаборатории.
Развитие молекулярно-генетических методов привело к появлению филогеографии, возможности соотнести генетическую дифференциацию таксона с изменениями физико-географических параметров в ходе геологической истории. Появление палеогеномики открывает возможность по сути дела напрямую сопоставлять современное генетическое разнообразие с генетическими параметрами животных, существовавших в прошлом, по крайней мере, в масштабах десятков тысяч лет.
Основой и специфической особенностью исследований, проводимых в лаборатории, является синтез палеонтологического и неонтологического подходов на базе современных представлений о микро- и макроэволюции как основе формирования эволюционного учения. Важным в этой связи является определение границ применимости актуализма как методологической основы такого синтеза».

Бородин Александр Васильевич – доктор биологических наук, заведующий лабораторией филогенетики и биохронологии Института экологии растений и животных (ИЭРиЖ УрО РАН) (г. Екатеринбург)
В настоящее время в лаборатории активно развиваются 4 основных направления: 1) исследование эволюционных (филогенетических) связей между группами организмов (видами, популяциями), реконструируемых на разных уровнях организации – морфологическом, цитогенетическом, молекулярном; 2) исследование процессов во времени, протекающих в биологических системах разного уровня от организменного до экосистемного во временных масштабах от актуального до геологического; 3) исследование экологических основ эволюционных процессов; 4) анализ генетического разнообразия и филогеографической структуры модельных видов позвоночных Северной Евразии.
На текущий момент одним из важных направлений работы лаборатории являются исследования в рамках гранта РФФИ "Биоразнообразие и генофонд фауны субарктических широт Евразии: глобальные и региональные факторы формирования и перспективы сохранения" под руководством Александра Бородина. В основе лежит опыт предыдущей многолетней работы, но с последующим выходом на новый методико-методологический уровень. Проект рассчитан на три года (2019-2021 гг.). В составе коллектива не только сотрудники лаборатории ИЭРиЖ УрО РАН, но и представители Института промышленной экологии РАН и Уральского федерального университета.
Как подчеркивает Александр Бородин, «традиционно изучение биоразнообразия проводят в рамках одного из подходов – либо палеонтологического, либо неонтологического, причем важнейшей задачей при этом является детальное описание таксономического состава фауны того или иного региона либо моделирование ниш отдельных видов. В данном проекте мы используем принципиально иной подход, направленный не на максимально полное описание современного биоразнообразия, а на выявление факторов, его обусловливающих. Для этого мы используем традиционные палеоэкологические подходы с выделением индикаторных видов, развиваемые, в том числе, и сотрудниками лаборатории, но применяем их к современным видам фауны субарктических широт. Перспективность данного подхода для реконструкции фаунистических связей в пределах средних широт Евразии показана нами ранее.
В качестве детерминант биоразнообразия фауны на глобальном уровне мы рассматриваем климат и факторы, его определяющие. Проект предполагает исследование исторических корней биоразнообразия современной субарктической фауны, основанное на сопоставлении динамики ареалов в историческом и геологическом масштабах времени с учетом цикличности действия климатогенных факторов и региональных геологических событий.
Для модельных видов современных зональных, интра- и азональных наземных местообитаний, различающихся по миграционным стратегиям, будет проведен комплексный анализ современного генетического разнообразия, описана и продатирована пространственно-временная динамика распространения в историческом и геологическом масштабах времени. Это позволит установить, как отвечают представители фауны на климатические изменения – индивидуально для каждого вида, либо могут быть выделены общие тенденции в зависимости от а) типа используемых местообитаний, б) характера изменения ареала во времени и пространстве, в) длительности эволюционной истории вида.
Совместно с нашими коллегами из Уральского федерального университета в ходе выполнения проекта будет использована наиболее совершенная модель долгопериодической эволюции орбиты Земли La2010, применяемая для калибровки геологических шкал времени, а также для оценки инсоляции поверхности Земли на интервалах времени в 50 млн. лет, как в прошлое, так и в будущее. Данная модель позволяет детально исследовать вариации инсоляции с высоким временным разрешением. Все это даст возможность сопоставить выявленные центры генетического и морфологического разнообразия модельных видов, времени и масштабов внутри и межвидовой дифференциации на фоне изменений видового состава и структуры региональных фаун севера Евразии с геологическими событиями и моделью инсоляции».
Александр Бородин ведет работы по описанию четвертичной истории зональных фаунистических комплексов Западной Сибири и предложил свой подход для межзональных биостратиграфических и биохронологических корреляций фаун плейстоцена в центральной части Северной Евразии, который также применим для фаун эоплейстоценового возраста. «В данном случае речь идет о необходимости формализации описания исходных данных палеонтологической летописи и унификации методов их сравнения. Формализация критериев корреляции геологических отложений является неизбежной тенденцией в исследованиях четвертичного периода (последние 2.6 млн. лет). На региональном уровне эта задача может быть успешно решена, но проблемы возникают, когда мы пытаемся установить синхронность процессов, протекавших в экосистемах географически удаленных регионов, для которых условия природной среды в одно и то же время были различными.
Надо еще раз подчеркнуть, что в наших исследованиях мы используем как данные о современных видах, ныне живущих (или только что живших) организмах, так и данные палеонтологической летописи. Но если первые относятся к биологической реальности, то вторые, ископаемые остатки когда-то живших организмов – к геологической. Когда мы говорим об истории становления современных видов, современных экосистем, мы подразумеваем, что можем выстроить некую последовательность событий, происходивших в биоте на протяжении геологической истории как отдельных регионов, так и планеты Земля в целом. А для каждого геологического этапа мы можем (или по крайней мере пытаемся) оценить изменчивость этих когда-то существовавших биологических объектов в пространстве. И в этом случае мы уже решаем задачи не биологии и экологии, а биостратиграфических корреляций, формализуемых в соответствующих разделах геохронологической шкалы.
Для каждого отдельного региона в рамках полноты его геологической летописи мы можем описать этапы становления биоты в этом регионе, описать состав и структуру фаун того или иного геохронологического уровня. В региональных геохронологических шкалах им присваивают имена собственные, связанные, как правило с региональными географическим названиями. И в разных регионах для одного и того же геохронологического уровня существуют (используются) собственные названия, для каждого региона есть своя собственная последовательность названий, за которыми стоят различия в составе и структуре фаун, различия эволюционного уровня таксонов, их формирующих. И сравнение, корреляция биостратиграфической шкалы одного региона с другим происходит традиционно также на этом описательном уровне. С другой стороны, для всех очевидны различия в современной биоте разных регионов, обусловленные ландшафтно-климатическими различиями. Но градиент климатических параметров существовал и в прошлом, что, естественно, сказывалось на видовом составе и структуре фаун, существовавших в одно геологическое время в географически удаленных регионах. Причём физико-географические условия в то время могли принципиально отличаться от современных, а в составе фаун наряду с предковыми формами современных видов были и те, что не дожили до наших дней – мамонты, пещерные львы, шерстистые носороги. Соответственно, необходима формализация выделения как биохронологических уровней, так и пространственных корреляций».
По словам ученого, традиционным объектом для биостратиграфических и биохронологических исследований являются полевки (Arvicolinae, Cricetidae, Rodentia). Они характеризуются быстрой эволюцией, обильной ископаемой летописью и широким географическим распространением. Бородин развернуто конкретизирует, что «время появления и исчезновения ключевого таксона в филетических линиях полевок, терминальными видами в которых являются современные виды, широко используются в настоящее время в качестве ключевых биохронологических маркеров, как в Евразии, так и в Северной Америке. Предложенные А.С. Тесаковым и коллегами, работающими преимущественно в Европейской части России, биохроны были выделены по времени появления и исчезновения ключевого таксона (эволюционной стадии, характеризующейся определёнными морфологическими характеристиками) в филетических линиях полевок, терминальными видами в которых являются современные степные пеструшки, узкочерепные и водяные полевки. Эти биохроны хорошо применимы к степным или лесостепным фаунам и за пределами Европейской России. Однако для территорий, где на протяжении четвертичного периода была выражена широтная зональность: южные степные фауны переходили в фауны гиперборейного комплекса, в состав которого наряду со степняками входили виды, ассоциирующимися и с лесными, и с тундровыми биотопами, а фауны севера могли быть только лемминговые, прямое сопоставление невозможно. Поэтому необходимо дифференцировать различия в составе и структуре фаун, вызванные изменениями климатических характеристик во времени, от различий, связанных с широтным градиентом климатических условий в пределах одного хроносреза.
В настоящее время Западно-Сибирская равнина характеризуется ярко выраженной широтной зональностью, начиная от арктических пустынь на севере до степей на юге. Фауна тундры принципиально отличается от фауны степей и полупустынь, но в пределах равнины существует довольно плавный переход от одного биома к другому в градиенте север-юг: тундра – лесотундра – тайга – лесостепь – степь. На территории Западно-Сибирской равнины (включая Зауральский пенеплен) на всех этапах четвертичного периода была выражена широтная зональность. И нами были выявлены различия в зональных фаунах Урала и Западной Сибири в четвертичное время. Мы можем наблюдать динамические изменения в распространении с севера на юг тундроподобных на севере, гиперборейных (по терминологии Н.Г. Смирнова) или безаналоговых на месте центрально-таёжной зоны и степных фаун на каждом временном уровне. Наличие в составе фаун полевок, характер распространения которых может трактоваться как полизональный или интраполизональный, позволяет провести сопоставление широтных вариантов региональных фаун. На основании широтного распространения представителей филетических линий ключевых таксонов полевок для позднего плейстоцена были выделены пять широтных биохронологических зон (см. Рис. 1), в которых выражен переход от фаун, представленных только степными формами к фаунам, в составе которых предковые формы видов, обитающих в настоящее время только в тундре. Поскольку эти зоны связаны с ареалами ключевых таксонов, для их обозначения был использован индекс A (от “area” (лат.) – ареал). Нумерация проводится с севера на юг по градиенту увеличения видового разнообразия полевочьих. Названия широтных биохронологических зон соответствуют названиям терминальных родовых (подродовых) таксонов эволюционных линий полевок, используемых в качестве руководящих форм для относительной датировки (см. Рис.1). В пределах Западной Сибири этот подход позволил сопоставить географически удаленные районы, фауны которых в принципе не совпадали по видовому составу.
Это возможность проведения границ между базовыми биогеографическими зонами в геологическом прошлом, когда физико-географические условия принципиально отличались от современных, но это отнюдь не отменяет описание и изучение особенностей региональных и локальных фаун. Как для неонтологии есть представление о биомах как совокупности экосистем одной ландшафтно-климатической зоны и региональных различиях фаун в пределах одного биома. Предложенный нами подход в совокупности с биохронологическими зонами MQR А.С. Тесакова позволяет формализовать корректные межрегиональные корреляции фаун четвертичного периода Евразии (по крайней мере в пределах от современных степей до тундр). Подход основан на пересечении ареалов видов различных природных зон и использовании полевок с наиболее обширными ареалами в качестве связующих звеньев при сравнении фаун в широтном градиенте. Выявление синхронных эволюционных стадий морфологических характеристик полевок, распространенных в соседних зонах, позволяет сопоставлять фауны регионов с выраженным широтным градиентом условий среды. Таким образом, предложенный подход позволяет не только оценить эволюционный уровень (относительный возраст) фауны, но и получить информацию о характеристиках биомов, существовавших в конкретные интервалы времени».
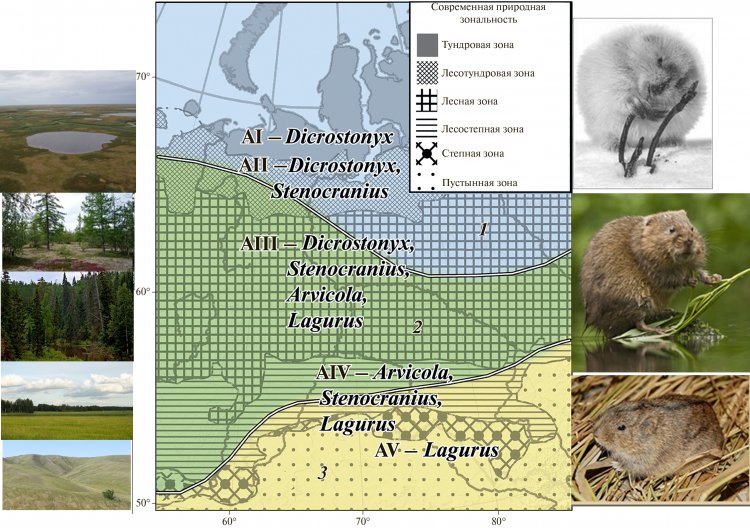
Рисунок 1. Границы распространения фаун мелких млекопитающих Урала и Западной Сибири в позднем плейстоцене. 1 – тундровые фауны, 2 – гиперборейные фауны, 3 – лесостепные и степные фауны; АI–АV – широтные биохронологические зоны, выделенные на основании совместного распространения руководящих форм полевок
На основе подхода, введенного Александром Бородиным для сопоставления фаун широтных зон плейстоцена, были получены результаты, которые по мнению ученого, «позволяют сформулировать основные ограничения для межзональных корреляций четвертичных фаун. Во-первых, это объективные факторы, влияющие на неравномерную сохранность комплексов ископаемых остатков в отложениях различных временных интервалов. Различия условий осадконакопления, а в горных регионах и изменения интенсивности тектонической активности, обусловливают неполноту ископаемой летописи. Во-вторых, это необходимость критического анализа тафономических факторов при выделении фаунистических комплексов. И, в-третьих, важно отметить необходимость совершенствования методов таксономической идентификации ископаемых остатков. Несмотря на имеющиеся ограничения, пять широтных биохронологических зон, выделенных на территории центральной части Северной Евразии, представляются удобной моделью для построения схем транс-евразийских фаунистических корреляций на протяжении разных этапов четвертичного периода».
Для учета широтной зональности Западной Сибири и Зауралья специалисты ИЭРиЖ УрО РАН предлагают подход к объединению межбиомных и межрегиональных корреляций фаунистических комплексов мелких млекопитающих, который основан на пересечении ареалов видов ключевых линий полёвок в различных природных зонах. Как отмечает Бородин, «выявление синхронных эволюционных стадий филетических линий полевок, распространенных в соседних зонах, позволяет сопоставить фауны регионов с выраженным широтным градиентом условий среды. Подход может быть потенциально использован для широтных зон всей континентальной Северной Евразии и применим для всего четвертичного периода».
Уральскими учеными были описаны этапы четвертичной истории зональных фаунистических комплексов на всей территории Урала (включая Предуралье и Зауралье) и Западно-Сибирской равнины – то, что административно называется Уральским регионом, поскольку географическое положение и особенности их геологической истории во время четвертичного периода определяют характер развития и видовое разнообразие современной фауны. По замечанию Александра Бородина, «в южных широтах палеонтологическая летопись представлена более полно, начиная с плиоцена, а чем дальше на север, тем более короткий временной интервал доступен для изучения (как правило не древнее позднего плейстоцена). Широтный климатический градиент был характерен для этой территории на протяжении всего четвертичного периода. Уникальность этого региона в том, что он являлся и является своеобразным биогеографическим перекрестком, через который шли миграции видов из Азии и Северной Америки в Европу, и наоборот. Регулировался этот процесс физико-географическими условиями в каждый конкретный момент геологической истории. Это могли быть покровные оледенения и сопутствующие им подпрудные озера, или морские трансгрессии Северного Ледовитого океана на Севере или Понто-Каспийского бассейна на юге, которые просто исключали часть суши для формирования наземных биомов и перекрывали пути миграций; в периоды регрессий, когда уровень океана понижался и шельф соединял Северную Америку и Евразию. Изменение параметров влажность/температура также влияло на изменение ареалов отдельных видов и формирование биомов. И, поскольку только площадь современной Западной Сибирской равнины составляет порядка 2,5 тыс. км², вклад этого региона в формирование биоты Евразии очевиден».

Ялковская Лидия Эдуардовна – кандидат биологических наук, старший научный сотрудник лаборатории филогенетики и биохронологии Института экологии растений и животных (ИЭРиЖ УрО РАН) (г. Екатеринбург
В рамках обсуждения исследований лаборатории были рассмотрены другие аспекты изучения биоразнообразия Северной Евразии. Сотрудники лаборатории Евгения Маркова и Лидия Ялковская поделились результатами своих научных разработок.
Исследования, которыми занимается Лидия Ялковская, дополняют существующие представления о внутривидовой генетической структуре и истории формирования современного ареала ряда широко распространенных видов.
Ученый отметила, для решения каких вопросов встает необходимость изучения различных филогрупп на территории Центральной Евразии: «В силу различных обстоятельств центральная часть Северной Евразии до сих пор остается "белым пятном" в генетических исследованиях многих широко распространенных видов Евразийского континента, в том числе, при анализе внутривидовой генетической структуры, распространения филогенетических линий, эволюционно-экологических факторов, обуславливающих генетическую дифференциацию и расселение внутривидовых группировок в пределах видового ареала. Тем не менее, без включения в анализ данных по таким крупным регионам как Урал, Западная Сибирь, Алтай, имеющим свои особенности геологической истории, отличающимся разнообразием климатических и физико-географических условий, предлагаемые гипотезы нельзя считать достаточно обоснованными».
По словам Ялковской, «наглядным примером важности анализа вклада физико-географических условий крупных регионов в формирование генетического разнообразия широко распространенных видов могут служить наши исследования налима из крупных речных систем центральной части Северной Евразии. До недавнего времени реконструкции эволюционной истории вида в Евразии проводили без учета генетических данных из богатейших по своим запасам вида рек Западной Сибири (Обь, Таз, Иртыш), а также крупнейшего в Европе Волжско-Камского речного бассейна. Сравнительный анализ полиморфизма митохондриального генома налима европейских, западносибирских и восточносибирских речных бассейнов показал, что реки Западной Сибири играли важную роль в сохранении вида в период последнего глобального похолодания, и в дальнейшем внесли значительный вклад в формирование генетического разнообразия налима Евразии и Северной Америки».
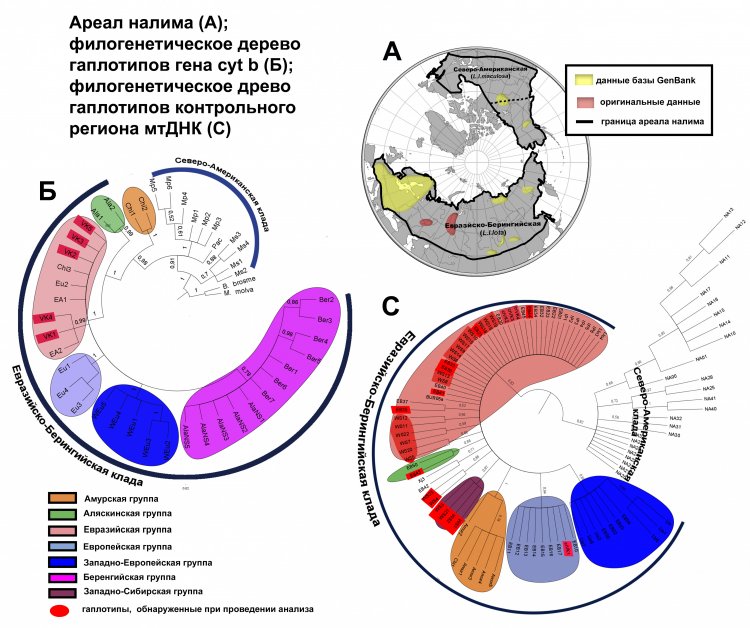
Рисунок 2. Положение исследуемых выборок налима в пределах ареала и результаты филогенетического анализа митохондриальных маркеров
В связи с актуальной на сегодняшний день проблемой сохранения биоразнообразия, важно исследование редких краснокнижных видов. Одно из исследований Лидии Ялковской было направлено на определение положения обыкновенной летяги Уральского региона в филогеографической структуре вида.
В чем специфика исследований редких видов, находящихся под охраной? Каково соотношение ареала и региона в формировании генетического разнообразия?
«Исследования редких видов сопряжены с рядом трудностей, связанных с получением материала для анализа, а также незначительным объемом данных других исследователей для сравнения получаемых результатов. Решением первой проблемы может стать использование музейных коллекций, как, например, в случае анализа филогеографии обыкновенной летяги нами были использованы коллекции Музея Института экологии растений и животных. Вторая проблема может являться даже плюсом, поскольку любые новые данные, полученные для редких видов, даже сами по себе представляют интерес. Наши исследования филогеографии обыкновенной летяги направлены на выявление роли Урала в формировании генетического разнообразия вида в пределах современного ареала. Оказалось, что в отличие от многих наземных грызунов, для обыкновенной летяги, ведущей древесный образ жизни, Уральские горы не являлись преградой в ходе постледникового расселения вида из Восточной в Западную Палеарктику. С помощью метода молекулярных часов нами были проведены расчеты времени происхождения вида и появления основных внутривидовых филогенетических линий. Полученные результаты сняли существующие противоречия между данными молекулярно-генетических исследований и палеонтологическими находками обыкновенной летяги в пещерах Алтая. – Уральский биолог указала на сложности, связанные с изучением природоохранных видов, и намеченные задачи дальнейшей работы. – В ближайшем будущем мы планируем продолжение исследований данного вида, как с использованием современных популяций, так и данных анализа палео-ДНК. Работа будет направлена на решение вопроса: "Почему обыкновенная летяга, успешно освоив территорию Дальнего Востока около 250-200 тыс. лет назад, в Западной Палеарктике появилась только в позднем голоцене? Каков уровень генетических отличий современной летяги и летяг, обитавших на Дальнем Востоке 40-10 тыс. лет назад и, по-видимому, являющихся предками популяций, обитающих в Европе в настоящее время?".
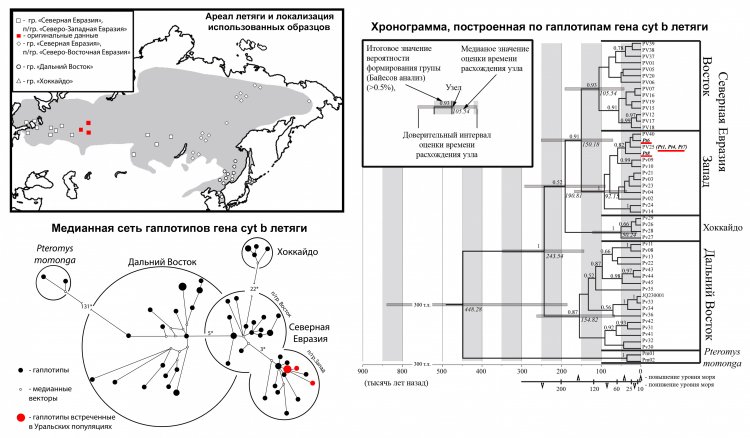
Рисунок 3. Ареал обыкновенной летяги, медианная сеть и хронограмма, построенные по гаплотипам гена cyt b
Кроме того, Лидия Ялковская исследовала как проявляется генетическая изменчивость на границах видового ареала на примере популяций желтогорлой мыши вблизи восточной границы ареала. Ученый разъяснила, что демонстрируют результаты филогенетического анализа и существует ли родство с популяциями из других частей видового ареала: «Одним из направлений исследований нашей лаборатории является анализ генетической изменчивости в популяциях широко распространенных видов, включая границы видовых ареалов. Так, нами проведены исследования желтогорлой мыши вблизи восточной границы распространения вида на Урале, в том числе в популяции, обитающей за пределами общепризнанных границ видового ареала. В результате анализа гена цитохрома b нами были не только подтверждены современные представления о генетической дифференциации вида и распространении основных филогенетических линий, но и реконструирована история появления изучаемых популяций вида на территории Урала. По-видимому, восточная граница ареала желтогорлой мыши сформировалась относительно недавно, в геологическом масштабе времени, в результате быстрого расселения одной из трех генетических линий, занимающей в настоящий момент территорию Балканского полуострова, Северной и Восточной Европы».
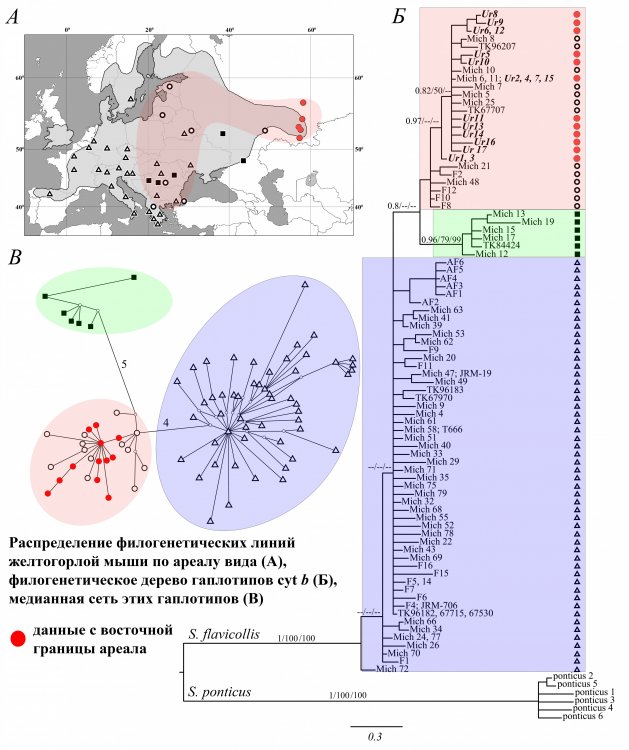
Рисунок 4. Локализация образцов желтогорлой мыши, включенных в анализ, и результаты филогенетических реконструкций
Как изучение формирования современной внутривидовой структуры поможет сохранению вида на фоне влияния глобальных климатических изменений и деятельности человека?
«Успешное существование вида обеспечивается его способностью адаптироваться к условиям среды обитания. Однако эти условия не являются константными, а подвержены постоянным изменениям, вызванными, в том числе, глобальными климатическими изменениями и хозяйственной деятельностью человека. Приспосабливаться к таким изменениям виду позволяет определенная генетическая структура, возникшая в ходе эволюции. Изучение формирования современной внутривидовой структуры с ретроспективной оценкой роли природных или антропогенных факторов позволяет спрогнозировать возможность сохранения вида при изменении условий среды обитания в будущем. Анализ генетической структуры вида необходим также для успешного восстановления численности вида в ходе интродукции, в частности при выборе популяций-доноров, генетические характеристики которых не должны значительно отличаться от аборигенных популяций и, в тоже время, обеспечивать уровень генетического разнообразия для дальнейшего выживания вида», – считает Лидия Ялковская.

Маркова Евгения Анатольевна – кандидат биологических наук, старший научный сотрудник лаборатории филогенетики и биохронологии Института экологии растений и животных (ИЭРиЖ УрО РАН) (г. Екатеринбург)
Евгения Маркова занимается исследованием экологических основ эволюционных процессов на примере мелких млекопитающих современной фауны и их предков различного эволюционного уровня. Одним из аспектов этих исследований является определение последствий критического сокращения численности популяций – так называемого популяционного бутылочного горлышка (population bottleneck) – для фенотипических признаков, определяющих приспособленность вида.
Какое место в эволюционной экологии и палеоэкологии занимают проблемы выявления фенотипических последствий критического сокращения численности популяций (бутылочного горлышка)?
«В классической эволюционной биологии со времен Чарльза Дарвина считалось, что маленькие изолированные группы способны выживать только искусственно, при поддержке человека, а в природе они неизбежно вымирают и признаки, накопившиеся в таких группах, стираются естественным отбором. Однако, бурное развитие генетики, особенно молекулярной генетики, на протяжении последних десятилетий, способствовало пониманию того, что в истории практически каждого биологического вида существовали периоды критического сокращения численности, сопровождавшиеся резким снижением генетического разнообразия. Именно благодаря заслугам генетики термин «бутылочное горлышко» вошел в обиход биологов-эволюционистов и стало понятно, что генетическая память о периодах критического сокращения численности сохраняется в генофонде вида. А если сохраняется генетическая память, то, как это отражается на признаках, определяющих приспособленность вида? И здесь нам на помощь приходят широко распространенные виды мелких млекопитающих. Благодаря обширным ареалам и быстрой смене поколений мелкие млекопитающие являются удобной моделью для изучения последствий критического сокращения численности и процессов, которые идут после ее восстановления из небольшого числа основателей в естественных условиях, – Евгения Маркова раскрыла суть понятия «бутылочное горлышко» и уточнила, какова роль редких признаков для изучения процессов трансформации ареала. – Мы показали, что морфологические аномалии, возникающие в результате близкородственного скрещивания, в природе не стираются полностью естественным отбором, а сохраняются в виде скрытых либо слабо выраженных признаков. Для эволюционной экологии это, безусловно, имеет значение, поскольку указывает на то, что условия, при которых формируются небольшие изолированные группы особей, являются не эволюционными тупиками, как это считалось ранее, а природными экспериментами, результатом которых отнюдь не всегда является вымирание. Полученные нами данные имеют значение и для палеоэкологии, то есть науки об экологических процессах и явлениях в уже исчезнувших экосистемах прошлых геологических эпох. Мы показали, что проявление одних и тех же редких признаков и аномалий зубной системы в географически удаленных популяциях одного вида может отражать процессы трансформации ареала под действием глобальных климатических изменений. Так, например, колебания уровня мирового океана могли приводить к периодическому формированию большого числа изолированных популяций на прибрежных территориях – и это мы можем проследить по особенностям зубной системы полевок и леммингов, живших, например, около 100000 лет назад».
Что представляет собой модель искусственного и естественного бутылочного горлышка и каким образом определяется, что популяция переживает бутылочное горлышко?
«Снижение численности и генетического разнообразия может происходить по естественным причинам либо в результате преднамеренных или непреднамеренных действий человека. Например, если изменился уровень мирового океана, и небольшая группа особей оказалась отрезана на вновь образовавшемся острове – это ситуация, соответствующая модели естественного бутылочного горлышка. А если же группа особей была интродуцирована человеком – то речь уже идет о ситуации искусственного бутылочного горлышка. В первом случае мы, как правило, имеем, дело с популяциями, существующими десятки тысяч лет, и ответить на вопрос о том, сопровождалась ли изоляция периодом критического сокращения численности, можем лишь реконструировав демографическую историю популяции по молекулярно-генетическим признакам. Во втором случае история популяции известна – мы изначально знаем такие важные моменты, как длительность изоляции, время интродукции, а иногда и число основателей – если вид был интродуцирован преднамеренно», – прояснила ученый.
Сравнивая современные виды млекопитающих и их предков различного эволюционного уровня, Евгения Маркова берет за объект признаки зубной системы.
Что выявляют закономерности изменчивости зубов у мелких млекопитающих, когда популяция переживает бутылочное горлышко? Как важные для выживания вида признаки зубов, строение жевательной поверхности отражают эти изменения?
«Мне кажется, что на современном этапе развития эволюционной биологии очень важно разрабатывать такие методы и подходы, которые позволяли бы изучать эволюцию в разных масштабах времени – от нескольких поколений до миллионов лет. К сожалению, на практике между работами палеонтологов, изучающих вымершие организмы, и теми, кто изучает микроэволюционные процессы в современных популяциях пересечений довольно мало. А ведь эволюция имеет под собой экологическую основу. Признаки зубной системы млекопитающих являются подходящим объектом для исследования экологических основ эволюционных процессов. С одной стороны, в строении зубов фиксируются этапы становления видов в палеонтологических масштабах времени и, что немаловажно – именно зубы хорошо сохраняются в ископаемом состоянии. С другой стороны, на современных видах мы можем решать задачи по выявлению экологических основ одонтологической изменчивости путем проверки гипотез о соотношении различных форм изменчивости и даже моделирования некоторых ситуаций. Например, мы рассматриваем лабораторные колонии грызунов как модель искусственного бутылочного горлышка с известным числом основателей, популяции видов с периодическими вспышками и депрессиями численности – как модель естественного бутылочного горлышка для локальных, зачастую эфемерных популяций, которые возникают во время пика численности на периферии ареала и т.д. В серии таких тематических исследований мы сопоставляем процессы, наблюдаемые на современных видах, с данными ископаемой летописи их предков различного эволюционного уровня. Это, в конечном итоге, позволяет нам переходить от описания закономерностей эволюции к выявлению механизмов, лежащих в ее основе.
Сопоставление с данными ископаемой летописи позволило, во-первых, подтвердить, что последствия близкородственного скрещивания могут быть обнаружены в ископаемой летописи, а во-вторых, показать, что в небольших изолированных группах за счет близкородственного скрещивания может происходить частичная утрата эволюционных достижений вида. Это возможно уже со второго поколения, а к девятому поколению, после преодоления инбредной депрессии, частота отклонений снижается, но могут появляться формы, рекапитулирующие признаки предковых форм, живших около миллиона лет назад (см. Рис.5), – подробно ответила Евгения Маркова. И подчеркнула вклад своих коллег и следование заложенным научным традициям – В этой связи мне хотелось бы отметить, что наш успех в данной области стал возможен благодаря тому, что в нашем Институте сохраняются традиции эволюционно-экологических исследований, заложенные С.С. Шварцем, А.В. Покровским, Э.А. Гилевой. Это касается и фундаментальной основы, и обширных материалов, накопленных под руководством этих выдающихся ученых и хранящихся в настоящее время в Музее ИЭРиЖ».
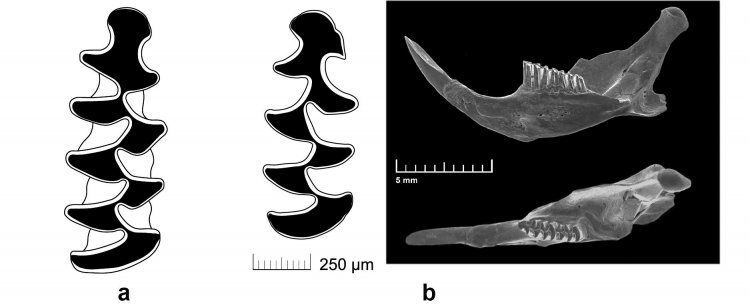
Рисунок 5. Упрощение первого нижнего зуба у обыкновенной полевки в девятом поколении лабораторной колонии до уровня предкового рода Allophaiomys, жившего около миллиона лет назад: а – контур жевательной поверхности типичного зуба современной обыкновенной полевки, b – контур зуба и фотография нижней челюсти лабораторной особи, не отличимой по строению первого нижнего зуба от предковой формы (по: Markova et al., 2020)
Как влияет территориальная удалённость и изолированность на признаки популяции показал анализ данных, собранных на Арктическом архипелаге Шпицберген. Ученые получили возможность исследовать степень и скорость морфологического расхождения, изучив размеры и морфологию зубов полевок, пойманных на Шпицбергене.
Каковы фенотипические различия популяций материкового вида и островного? Что показал анализ изменчивости зубов у островной популяции?
Евгения Маркова объясняет: «Одним из объектов наших исследований были популяции восточноевропейской полевки, сформировавшиеся в результате непреднамеренной интродукции далеко за пределы основной области распространения – на север Западной Сибири, Дальний Восток, Шпицберген. Считается, что полевки были завезены в эти отдаленные регионы в результате морских и железнодорожных перевозок в XX веке. Мы установили, что в тех случаях, когда интродукция сопровождается изоляцией, может происходить проявление скрытой фенотипической изменчивости и накопление редких особенностей в строении зубов. При этом не важно, происходит изоляция на острове за полярным кругом или же на материке в относительно мягком климате – важно, чтобы формировались условия изоляции на небольшой территории. На Шпицбергене такой ограниченной территорией является узкая прибрежная полоса на склонах под колониями морских птиц. Существование на ограниченной территории послужило причиной того, что эффект увеличения размеров, часто встречающийся у островных форм мелких млекопитающих, у восточноевропейской полевки на Шпицбергене не прослеживается. По размерам эти островные полевки не отличаются от своих материковых собратьев».
Тем не менее в природе достаточно часто складываются условия, способствующие формированию небольших изолированных популяций и благоприятствующие генетическому дрейфу.
Как выявленные факты о малочисленных локальных популяциях помогают решать проблему сохранения биоразнообразия?
«В ходе выполнения одного из проектов, поддержанных РФФИ, мы установили, что не только генетические признаки, но и особенности строения зубов могут служить маркерами того, что в прошлом предки данной особи прошли через стадию критического сокращения численности. В нескольких тематических исследованиях в рамках этого проекта мы также показали, что в природе небольшие изолированные популяции, сходные по ряду признаков с лабораторными колониями, которые создает человек, формируются в определенных ландшафтных условиях – в прибрежных зонах, на островах, в предгорьях. Во всех этих случаях мы наблюдаем повышение роли случайных факторов в формировании облика изолированной популяции. Благодаря эффекту основателя такие локальные популяции могут существенно отличаться друг от друга. Но общим в них является одно – проявление, а иногда и накопление одних и тех же редких признаков и аномалий в результате того, что близкородственное скрещивание снимает запреты на реализацию скрытой изменчивости. – Евгения Маркова выделила главные достижения проделанной работы. – В итоговой статье по проекту (QUATERNARY SCIENCE REVIEWS 228 (2020) 106045) мы обобщили значение этих результатов для тех, кто изучает ископаемую летопись мелких позвоночных за последние 2.6 млн. лет. Но полученные результаты потенциально могут быть использованы и в неонтологии, в частности для мониторинга современных популяций. Например, зубные признаки-маркеры близкородственного скрещивания могут быть информативны при изучении структуры популяций видов, чьи ареалы претерпевают быстрые изменения – сокращение или расширение. Введение разработанных методик в практику мониторинга природных популяций леммингов и полевок в перспективе может быть полезно для решения проблемы сохранения биоразнообразия в первую очередь арктических и субарктических экосистем, для которых эти мелкие млекопитающие являются ключевым звеном».
Итак, исследования лаборатории филогенетики и биохронологии ИЭРиЖ УрО РАН направлены на уточнение данных генофонда и выяснение истории фауны центральной части Северной Евразии, а также на реконструкцию условий, на фоне которых сформировалось современное биоразнообразие фауны Урала и Западной Сибири. Исследования генетической структуры и анализ одонтологических признаков популяций млекопитающих служат основой реконструкции демографической истории ключевых видов фауны и вместе с данными ископаемой летописи помогают выявлять фундаментальные закономерности микро- и макроэволюции, а также получать практические результаты, важные для разработки научно обоснованной стратегии охраны биологического разнообразия.
«Проблема сохранения биоразнообразия в условиях трансформации экосистем в ответ на изменения средовых характеристик актуальна в связи с риторикой вокруг возможных последствий глобальных климатических изменений и вклада в них антропогенной составляющей. Установленное в настоящее время повышение среднегодовых температур на поверхности Земли в высоких широтах Северного полушария проявляется наиболее остро. Несмотря на то что изменения поступления солнечной энергии в масштабах актуального времени пренебрежимо малы по сравнению с общим расчетным падением температур, в течение тысячелетий в высоких широтах Северного полушария отмечены биотические изменения, согласующиеся с температурными изменениями последних десятков лет. Данные о сокращении ареалов и/или численности типичных арктических видов и продвижение на север видов умеренных широт косвенно указывают на то, что ответ биоты на климатические изменения может быть прослежен в актуальном масштабе времени. В Субарктике эти изменения проявляются наиболее отчетливо, что позволяет рассматривать субарктические широты в качестве модельных для выяснения механизмов ответа биоты на изменения климата в различных масштабах времени – от актуального до геологического. – Руководитель проекта Александр Бородин дал оценку полученным результатам исследования и очертил круг предстоящих задач фундаментального и прикладного порядка. – Одним из приоритетных направлений деятельности лаборатории является исследование отклика биоты субарктических широт Северного полушария на действие циклических климатогенных факторов в актуальном, историческом и геологическом масштабах времени (с учетом региональных геологических событий). Эту задачу мы решаем на примере пойкилотермных (насекомых) и гомойотермных (мелких млекопитающих) представителей фауны с учетом их места в экосистемах – как существующих, так и уже исчезнувших, но реконструируемых с использованием комплексного палеоэкологического подхода. Практическое значение работы определяется возможностью разработки эффективных методов мониторинга и стратегий охраны видов наземной фауны в условиях естественной динамики ареалов. Сотрудничество с коллегами с кафедры астрономии, геодезии и мониторинга окружающей среды Уральского федерального университета позволяет нам рассматривать наши результаты в контексте астрономической теории климата, мы планируем их представить в форме, обеспечивающей возможность последующего использования для построения комплексных палеоклиматических моделей, учитывающих внутренние механизмы функционирования климатической системы Земли в применении к центральной части Северной Евразии».
























{kind=link}