

В центре внимания ученых из КарНЦ РАН образование уникальной древесины карельской березы, известной своим неповторимым декоративным узором. Изучая молекулярно-генетические и физиологические механизмы, биологи открывают возможности регулирования процесса образования древесины, или ксилогенеза. Полученные данные могут быть использованы при разработке рекомендаций для выращивания карельской березы и других древесных растений
Древесина карельской березы по рисунку напоминает мрамор и по праву считается самой декоративной древесиной Севера. Важную роль в формировании ее рисунка играют извилистые очертания годичных колец, темноокрашенные включения различных очертаний и перламутровый блеск. Уже более тридцати лет специалисты из Института леса Карельского научного центра РАН изучают этот уникальный феномен – образование узорчатой древесины карельской березы, выявляют генетическую предрасположенность к ее появлению и исследуют влияние условий среды на степень развития узора древесины.
Наталия Алексеевна Галибина – доктор биологических наук, заместитель директора по научной работе Института леса КарНЦ РАН, руководитель лаборатории биотехнологии растений Отдела комплексных научных исследований КарНЦ РАН (г. Петрозаводск) – рассказала об исследованиях процесса ксилогенеза (образования древесины), в том числе, и аномальной (узорчатой) древесины карельской березы.

На фото – Наталия Алексеевна Галибина – доктор биологических наук, заместитель директора по научной работе Института леса КарНЦ РАН, руководитель лаборатории биотехнологии растений Отдела комплексных научных исследований КарНЦ РАН (г. Петрозаводск)
Наталия Галибина подробно изложила, почему важно изучение механизмов ксилогенеза древесных растений:
«Древесные растения являются основными продуцентами в лесных экосистемах, а древесина представляет собой важный возобновляемый источник органического углерода. Ускорение роста и улучшение качества древесины, ее технологических свойств – основные задачи современной лесной биотехнологии. Для древесины (ксилемы) характерен широкий спектр составляющих ее структурных элементов ‒ это сосуды (функция водопроведения), трахеиды (основная функция механическая, но участвуют в проведении воды), волокна (механическая функция), клетки паренхимы, образующие радиальные лучи (функции транспорта и запасания питательных веществ). Формирование древесины происходит за счет притока фотоассимилятов из листьев по флоэме. Флоэма (нисходящий транспорт продуктов фотосинтеза) и ксилема (восходящий транспорт воды и растворенных в ней минеральных веществ) образуют проводящие ткани древесного растения. Обе ткани (ксилема и флоэма) образуются в результате деятельности латеральной (боковой) образовательной ткани или меристемы ‒ камбия. Меристемы представляют собой ниши, в которых находятся стволовые клетки, участвующие в процессах роста и регенерации растительного организма. Стволовые клетки – это недифференцированные клетки, способные к симметричному и ассиметричному делению и дифференцировке в разные типы клеток и могут дать начало не только определённой ткани, органу, но и целому растительному организму. При делении такой клетки одна из образовавшихся дочерних клеток остается стволовой, а другая становится предшественницей структурных элементов флоэмы (расположена наружу от стволовой клетки), или элементов ксилемы (расположена внутрь от стволовой клетки). Такие стволовые клетки, которые могут производить клетки и флоэмы, и ксилемы, называются бифасциальными (двухсторонними).
Этапы дифференциации клеток камбия в структурные элементы ксилемы включают рост клеток растяжением, формирование вторичной клеточной стенки, а в случае волокон, сосудов, трахеид ‒ программируемую клеточную смерть (ПКС). В отличие от слабодифференцированных паренхимных клеток, это мертвые клетки, которые состоят только из клеточных стенок. Клеточные оболочки в древесных растениях составляют до 90% от сухой массы. По сравнению с первичной клеточной стенкой, вторичная клеточная оболочка много толще и составляет большую часть целлюлозной биомассы, которая в том числе служит возобновляемым ресурсом для производства биотоплива. Структурные элементы древесины отличаются между собой по составу компонентов их клеточных стенок (целлюлоза, гемицеллюлозы, пектины, лигнин, белки). Соотношение структурных элементов в древесине, состав их клеточных стенок определяют ее физико-механические, технологические, топливные и декоративные свойства. Это очень краткий рассказ о процессах формирования древесины, который необходим для понимания дальнейшего повествования. Важно понять, что поиск путей эффективного управления ксилогенезом (процессом формирования древесины) необходим как с точки зрения повышения продуктивности растений, так и для получения древесины с заданными свойствами».
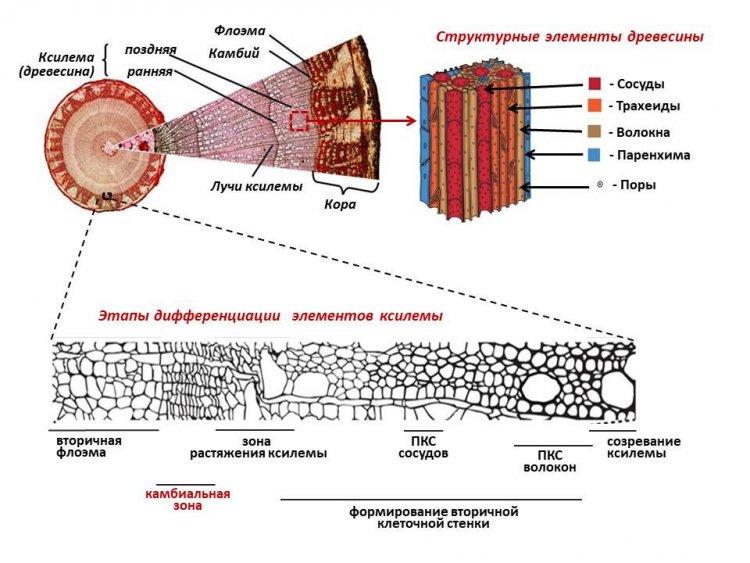
Рисунок 1. Структурные элементы древесины и схематическое отображение основных этапов ее образования
В последнее десятилетие впечатляющие достижения в клеточной, молекулярной и структурной биологии, как отмечает Галибина, «привели к интегрированному взгляду на образование древесины, начиная с деления клеток в камбии, роста клеток и утолщения клеточных стенок, вплоть до запрограммированной клеточной смерти. Широко распространенными экспериментальными моделями для изучения физиолого-биохимических и молекулярно-генетических механизмов регуляции процесса ксилогенеза является клеточная культура Zinnia elegans L. (Цинния изящная – однолетнее травянистое растение), у которой при выращивании в среде с гормонами (ауксином и цитокинином) клетки трансдифференцируются в трахеальные элементы, и Arabidopsis thaliana (Резушка Таля – модельный организм для изучения генетики и биологии развития растений), у которого типы сосудистых клеток вторичной ксилемы аналогичны типам тополя, но только меньше по размеру и лишены клеток лучевой паренхимы. Понятно, что с травянистыми растениями работать легче, но недавно появились работы, в которых уже на модельном древесном растении – тополе изучают механизмы ксилогенеза».
Во всем мире растет интерес к изучению молекулярных механизмов, регулирующих формирование тканей ствола древесных растений. Интерес «подогревается» и тем, что именно древесные растения играют значительную роль в сохранении климата, «консервируя» углекислоту. Для понимания этих механизмов создают конструкции, в которых одни гены «выключают», а другие заставляют работать активнее (сверхэкспрессируют).
«Особенностью нашего исследования является изучение ксило- и флоэмогенеза у древесных растений в ходе отклонений от нормального роста и развития проводящих тканей. Изучение отклонений от нормы позволяет нам глубже и всесторонней постигнуть механизмы регуляции нормального процесса», – комментирует Наталия Галибина, – Сама природа подарила нам уникальный экспериментальный объект – карельскую березу, в стволе которой одновременно протекают два процесса – нормальный и аномальный ксилогенез. Карельская (Betula pendula var. carelica) – это форма березы повислой (Betula pendula var. pendula), у которой формируется только типичная для вида прямослойная древесина. Деревья карельской березы различаются между собой по времени начала и моделям развития узорчатой древесины в онтогенезе. Бывает и такое, что узорчатая древесина формируется только на одной стороне ствола. Представьте – в рамках одного ствола мы можем исследовать "нормальные и аномальные" сценарии роста и развития проводящих тканей, нивелируя тем самым влияние факторов среды».

Рисунок 2. Деревья карельской березы сильно различаются между собой по времени начала и моделям развития узорчатой древесины в онтогенезе
Чем обоснован выбор темы научного исследования и какова определяющая роль научного наставника Наталия Галибина пояснила:
«К изучению механизмов формирования аномальной древесины я подключилась в 2006 году. Это направление исследований возглавляет доктор биологических наук, Людмила Людвиговна Новицкая. Людмила Людвиговна – признанный специалист в области анатомии и физиологии древесных растений, под ее руководством ведутся работы по изучению структурно-функциональных особенностей формирующихся тканей ствола древесных растений, а также регуляторных взаимоотношений флоэмы, камбия и ксилемы в ходе онтогенетического развития растений. Область моих интересов, в большей степени, сосредоточенна на изучении физиолого-биохимических и молекулярно-генетических механизмов функционирования стволовых клеток камбия, как в норме, так и при формировании структурных аномалий».

На фото – Людмила Людвиговна Новицкая – доктор биологических наук, главный научный сотрудник лаборатории физиологии и цитологии древесных растений Института леса КарНЦ РАН (г. Петрозаводск)
«Можно уверенно утверждать, что на сегодняшний день в Институте леса сложился творческий и, можно сказать, молодежный коллектив, включающий специалистов разных направлений, что позволяет всесторонне подходить к изучению механизмов ксилогенеза. Исследованием анатомо-морфологических особенностей структурных аномалий древесных растений занимаются кандидат биологических наук, старший научный сотрудник Надежда Николаевна Николаева и научный сотрудник Татьяна Владимировна Тарелкина. Кандидат биологических наук, старший научный сотрудник Юлия Леонидовна Мощенская и научный сотрудник Ксения Михайловна Никерова исследуют различные метаболические пути, протекающие при формировании нормальной и аномальной древесины, изучают гены, регулирующие эти процессы, а также выявляют физиолого-биохимические маркеры разных сценариев ксилогенеза у древесных растений. Полевые исследования, в которых, как правило, принимают участие все члены коллектива, приурочены к активному росту растений (май-июль) и включают описание объектов и условий их произрастания, постановку экспериментов, отбор растительного материала. Органы и ткани древесных растений замораживают в жидком азоте и хранят в низкотемпературных морозильных камерах при -80оС до проведения анатомических, физиолого-биохимических и молекулярно-генетических анализов. Основную часть этой очень важной работы выполняют ведущие биологи лаборатории физиологии и цитологии древесных растений ИЛ КарНЦ РАН: Подгорная Марина Николаевна, Софронова Ирина Николаевна, Семенова Людмила Игоревна, Иванова Диана Сергеевна», –Галибина отметила значимый вклад своих коллег в работу по изучению механизмов ксилогенеза.

На фото – Мощенская Юлия Леонидовна – кандидат биологических наук, старший научный сотрудник лаборатории физиологии и цитологии древесных растений; Никерова Ксения Михайловна – научный сотрудник, руководитель лаборатории аналитическая; Николаева Надежда Николаевна – кандидат биологических наук, старший научный сотрудник лаборатории физиологии и цитологии древесных растений; Тарелкина Татьяна Владимировна – научный сотрудник лаборатории физиологии и цитологии древесных растений Института леса КарНЦ РАН (г. Петрозаводск).

На фото – ведущий биолог Подгорная Марина Николаевна, ведущий биолог Софронова Ирина Николаевна, ведущий биолог Иванова Диана Сергеевна, ведущий биолог Семенова Людмила Игоревна лаборатории физиологии и цитологии древесных растений Института леса КарНЦ РАН (г. Петрозаводск).
Как происходит образование уникальной текстуры древесины карельской березы? Какие физиолого-биохимические особенности лежат в основе формирования узорчатой древесины?
«Оригинальная текстура древесины карельской березы формируется в результате отклонений в деятельности латеральной меристемы – камбия. В зонах развития структурных аномалий не запускается программа гибели клеток, приводящая к формированию сосудов и волокон ксилемы и ситовидных элементов флоэмы. Камбиальные производные дифференцируются в клетки запасающей паренхимы, которые накапливают большие количества запасных веществ. Среди запасных веществ много фенольных компонентов, которые на спиле проявляются в виде темноокрашенных включений. При этом образующиеся волокна и сосуды имеют неправильную искривленную форму, обеспечивая древесине карельской березы свилеватость и перламутровый блеск. В аномальных участках можно выделить два основных отличия: (1) увеличение количества делений в камбиальной ткани в целом и (2) изменение ориентации деления клеток камбия (они начинают делиться главным образом поперечными и антиклинальными перегородками вместо переклинальных, доминирующих при образовании нормальной древесины березы)», – объяснила ученый
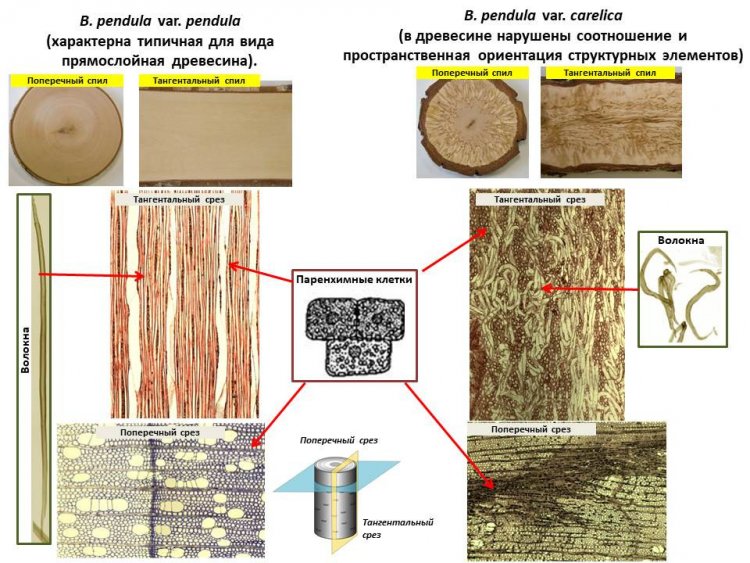
Рисунок 3. Макро- и микроструктура древесины двух форм березы повислой
Чтобы понять, как происходит формирование оригинальной текстуры древесины у карельской березы, ученые Института леса КарНЦ РАН изучают метаболические процессы, происходящие в местах формирования структурных аномалий.
Наталия Галибина сообщила об особенностях обменных процессов, наблюдаемых при формировании узорчатой древесины карельской березы:
«Нами впервые было показано, что именно сахароза (основная транспортная форма ассимилятов у березы повислой) играет важную роль в формировании структурных аномалий у карельской березы. Сахароза – химически инертна, для того чтобы она вступила в метаболические процессы необходимы ферменты, которые катализируют ее превращения. Утилизация сахарозы происходит при участии ферментов ее гидролиза – сахарозосинтетазы и инвертазы. Сахарозосинтаза в присутствии уридиндифосфата (УДФ) обратимо расщепляет сахарозу на уридиндифосфатглюкозу (УДФГ) и фруктозу (сахароза + УДФ ↔ УДФ-глюкоза + фруктоза). Несмотря на двойную направленность сахарозосинтазной реакции, в акцепторных тканях, к каким относится камбиальная зона, сахарозосинтаза функционирует, в основном, в направлении распада сахарозы. В ходе ксилогенеза сахарозосинтаза активно включается в метаболизм клеток на этапе отложения вторичной клеточной стенки, регулируя количество УДФ-глюкозы, идущей на синтез микрофибрилл целлюлозы. Данная стадия является одной из наиболее важных, поскольку клеточные стенки составляют основную часть древесной биомассы. Инвертаза катализирует реакцию гидролиза сахарозы до глюкозы и фруктозы (сахароза + Н2О → глюкоза + фруктоза). Гексозы, образующиеся в ходе инвертазного расщепления сахарозы, могут расходоваться на дыхание, синтез крахмала и компонентов клеточных стенок (целлюлоза, гемицеллюлозы, лигнин и др.), а также выступать в роли сигнальных молекул. Соотношение активностей сахарозосинтазы и разных форм инвертазы определяет преимущественное включение расщепляемой ими сахарозы в те или иные метаболические пути и, как следствие, оказывает влияние на направление дифференциации производных камбия».
Как подчеркивает ученый, специалистами КарНЦ РАН «впервые были выявлены физиолого-биохимические маркеры разных сценариев ксилогенеза (формирование нормальной и аномальной древесины). Формирование нормальной по строению древесины обычной березы, в составе которой преобладают сосуды и волокна, происходит на фоне высокой активности сахарозосинтазы и сопровождается активным синтезом структурных компонентов клеточных стенок. В этих тканях наблюдается высокий уровень экспрессии генов, кодирующих сахарозосинтазу (Sus1,2) и целлюлозосинтазу (Ces3,7,8). Включение сахарозы через сахарозосинтазный путь в синтез целлюлозы приводит к необратимому выводу ее из обмена веществ. В зонах структурных аномалий узорчатой древесины, где преобладает дифференциация производных камбия в паренхимные клетки, активность сахарозосинтазы низкая (снижение экспрессии генов Sus1,2) и уменьшается содержание целлюлозы в ксилеме (уровень экспрессии генов, кодирующих целлюлозосинтазу (Ces3,7,8), снижен). В этом случае метаболизация притекающей по флоэме сахарозы происходит с участием апопластной инвертазы. При этом уменьшение отношения активности сахарозосинтаза / апопластная инвертаза коррелирует со степенью развития структурных аномалий в камбиальной зоне березы повислой.
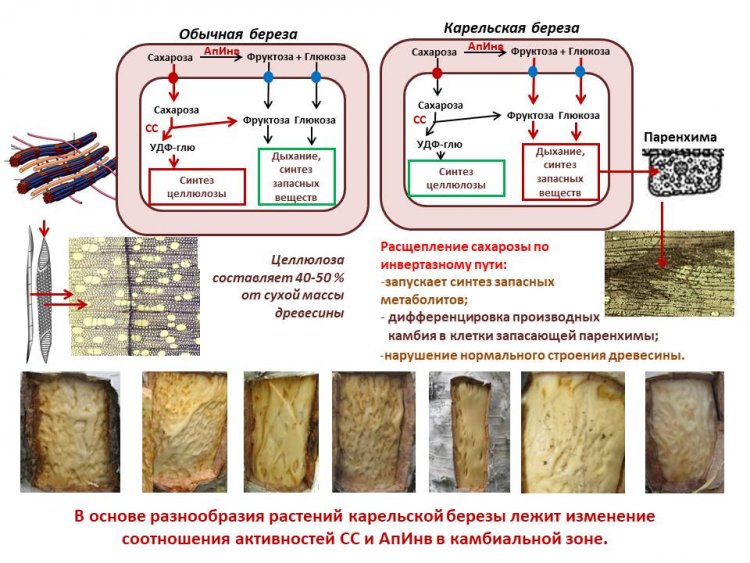
Рисунок 4. Схематическое изображение путей утилизации сахарозы в клетках камбиальной зоны при формировании нормальной и аномальной древесины
Высокая активность апопластной инвертазы в узорчатых участках ствола сопровождается интенсивным образованием гексоз, использование которых в реакциях энергетического /пластического обмена может приводить к генерации активных форм кислорода и возрастанию фенольных компонентов, оказывая тем самым влияние на активность ферментов антиоксидантной системы (АОС). В камбиальной зоне у двух форм березы повислой мы исследуем активность следующих ферментов АОС: супероксиддисмутазы, переводящей супероксид-радикал в перекись водорода, каталазы и пероксидазы, участвующих в утилизации перекиси водорода, и полифенолоксидазы, принимающей участие в процессах синтеза лигнина. Мы показали, что формирование узорчатой структуры древесины у карельской березы сопровождается повышением активности ферментов АОС, которые можно использовать в качестве биохимических маркеров узорчатости древесины».
Развитие структурных аномалий узорчатой древесины карельской березы обусловлено взаимодействием многих процессов. В основе нарушения развития камбиальных производных лежат сложные молекулярно-генетические механизмы.
«Один из них – это инактивация гормона ауксина (ИУК) через образование его коньюгата ИУК-глюкоза, нарушение транспорта гормона. У карельской березы важными показателями аномального развития ксилемы являются уменьшение в ней доли сосудов, нарушение их пространственной ориентации и усиленная паренхиматизация ткани. Индуктором дифференцировки сосудов служит ауксин, направление его потоков определяет пространственную ориентацию сосудов. Мы установили, что появление структурных аномалий древесины у карельской березы связано со сверхэкспрессией гена, кодирующего фермент ИУК-глюкозасинтаза, который катализирует реакцию инактивации ауксина путем синтеза его коньюгата ИУК-глюкоза. Показано, что синтез ИУК-глюкоза через цепь биохимических реакций может быть связан с гексозами, которые образуются в апопласте в результате расщепления сахарозы инвертазой клеточных стенок. При этом изменятся локализация на плазмалемме PIN белков, участвующих в транспорте ИУК, результатом чего становится изменение направления транспорта гормона. У карельской березы это находит отражение в нарушенной ориентации сосудов ксилемы. Мы полагаем, что появление "вихрей" и "струй" в узорчатой древесине карельской березы обусловлено именно этими процессами», – разъяснила Наталья Галибина.
С какого возраста и каковы начальные признаки проявления узорчатости карельской березы и в чем причина их изменений?
«Видимые появление признаков узорчатой древесины у карельской березы обычно можно наблюдать в возрасте 4-5 и 7-10 лет, в зависимости от условий произрастания. Наши исследования показали, что уже на ранних этапах развития (до появления видимых отличий) у карельской березы имеют место изменения путей утилизации сахарозы, которые во взрослом состоянии являются индикаторами нарушений в камбиальной деятельности. Показано, что активность ферментов АОС может являться тест-признаком для диагностики аномального ксилогенеза у карельской березы уже на самых ранних этапах онтогенеза», – конкретизирует ученый, –
Мы исследуем деревья обычной (типичная для вида прямослойная древесина) и карельской (узорчатая древесина ствола) березы повислой разного возраста – это и взрослые (40-45-летние) растения, 6-14-летние растения, у которых начинает формироваться узорчатая древесина, а также 2-4 летние сеянцы, выращенные из семян обычной и карельской березы. В лабораторных условиях в камерах при контролируемых температуре и освещенности мы выращиваем сеянцы обычной и карельской березы».
По замечанию Галибиной, «при любом варианте скрещивания родительских форм в потомстве карельской березы появляются особи, как с узорчатой, так и безузорчатой текстурой древесины. Среди опытных деревьев карельской березы мы подбираем экземпляры с высокой степенью узорчатости древесины и растения, у которых признаки аномального строения древесины отсутствовали (безузорчатые растения)».
Большинство исследованных учеными растений произрастает на Агробиологической станции Карельского научного центра, а также на лесосеменных плантациях на территории Республики Карелия. По словам Галибиной, «ареал карельской березы прерывистый. В пределах области естественного распространения она не образует чистых по составу древостоев, а встречается одиночно или группами в смешанных насаждениях с другими лиственными, а иногда хвойными видами деревьев. Диапазон экологических условий, в которых произрастает карельская береза, довольно узок и определяется хорошим освещением, температурным режимом, способствующим интенсивному оттоку сахарозы из листьев, нормальным увлажнением. Почвенные условия и потребности в питательных веществах, в частности соотношение азота и фосфора, непосредственно влияют на рост растений, углеводный обмен, активность ферментов, и как следствие, на формирование древесного узора. Анализ почвы в Кондопожском, Пряжинском и Медвежьегорском районах Карелии показал, что в этих лесах растут карельские березы с различной интенсивностью рисунка».

На фото – Никерова Ксения Михайловна, Подгорная Марина Николаевна, Галибина Наталия Алексеевна, Воробьев Виталий Владимирович, Софронова Ирина Николаевна, Николаева Надежда Николаевна, Мощенская Юлия Леонидовна на лесосеменной плантации карельской березы в Медвежьегорском районе
Как почвенные условия влияют на формирование и качество древесины карельской березы?
«В результате изучения мест естественного распространения карельской березы и анализа характеристик почв в пределах ее ареала Л.Л. Новицкой было высказано предположение, что карельская береза не распространяется как в области очень бедных, так и относительно богатых почв. Наши исследования показали, что большое влияние на проявление признака узорчатости древесины у карельской березы оказывает доступность подвижного азота в почве. В условиях нормального обеспечения азотом формирование узора идет на фоне интенсивного роста (высокие активности сахарозосинтазы и апопластной инвертазы в ксилеме и активность апопластной инвертазы во флоэме). Некоторое снижение плодородия почвы приводит к снижению активности сахарозосинтазы на фоне возрастания активности апопластной инвертазы и сопровождается увеличением количества темноокрашенных включений паренхимы в древесине. При этом морфологические показатели растений не изменяются. При сильном дефиците подвижного азота в почве усиливается конкуренция за фотоассимиляты между формированием узорчатой древесины (процесс связан с усиленной метаболизацией сахарозы по апопластному пути во флоэме) и радиальным ростом ствола (использование сахарозы в ксилеме на формирование клеточных стенок).
Результаты исследования могут быть использованы для объяснения, почему ареал карельской березы не включает плодородные почвы. Под воздействием высокого уровня азотного питания (нитратов) в ксилеме карельской березы снижается метаболизация сахарозы по сахарозосинтазному и инвертазному пути. Первое ведет к уменьшению приростов ксилемы, второе – к снижению в ней содержания паренхимы, т.е. подавлению формирования узорчатой текстуры древесины. В этих условиях сахароза, поступающая из листьев, расходуется во флоэме. Здесь с участием апопластной инвертазы она интенсивно используется в реакциях запасного метаболизма, что поддерживает жизнедеятельность существующих и образование новых клеток запасающей паренхимы. Поэтому отличительной особенностью берез, произрастающих на плодородных почвах, становится толстая кора, а максимальным выражением этих тенденций – появление грубокорых форм березы», – ответила Наталья Галибина.
Таким образом, всесторонние, в том числе, молекулярно-генетические исследования древесных растений учеными КарНЦ РАН представили новый взгляд на механизмы, лежащие в основе формирования узорчатой древесины.
По словам карельского ученого, «многолетние исследования членами коллектива насаждений карельской березы в пределах ее естественного ареала и в лесных культурах на территории Карелии позволили получить данные, характеризующие анатомо-морфологические и физиолого-биохимические особенности деревьев с разной степенью проявления аномальности строения древесины. Получен патент на полезную модель «Способ диагностики узорчатой текстуры древесины карельской березы» (Авторами Н.А. Галибиной и К.М. Никеровой). Члены коллектива разработали ряд экспериментальных подходов, целью которых было нарушение у растений обычной березы транспорта сахарозы и, соответственно, создание зон ее избыточного содержания. Получен патент на полезную модель «Способ выращивания древесины с декоративной текстурой» (Авторы Л.Л. Новицкая, Ф.В. Кушнир). Высказана гипотеза о возможной роли повышенного содержания сахарозы в формировании аномальной древесины у карельской березы (публикации в высокорейтинговых журналах).
Благодаря комплексному методическому подходу, включающему выбор уникального объекта, разработку экспериментальных подходов для получения слоев ксилемы и флоэмы с преобладанием в них одного из типов структурных элементов, использованию физиолого-биохимических, молекулярно-генетических методов, создана основа для разработки способов управления ксилогенезом с целью увеличения выхода биомассы древесины и выращивания древесного материала с заданными свойствами. Результаты, полученные в ходе изучения уникального объекта – карельской березы– мы используем в качестве фундаментальной основы для выявления механизмов ксилогенеза у основных лесообразующих пород (сосны и ели)».























{kind=link}